
14 Chapter 14: Sex-determination Systems
Joshua Reid and Lisa Limeri
Learning Objectives
At the end of this section, you will be able to:
- Explain how sex chromosomes contribute to sex determination in some organisms.
- Describe the variation that exists within genetic sex determination (GSD) mechanisms.
- Explain the function of aromatase in temperature-dependent sex determination (TSD) mechanisms.
- Compare genetic sex determination (GSD) mechanisms with temperature-dependent sex determination (TSD) mechanisms.
Introduction
There is great variety in the ways sex of individuals is determined in biology. Some systems are based on genetics, while others are based on environmental factors. Individuals of some species change their sex during their lives, alternating between male and female. If the individual is female first, it is termed protogyny or “first female;” if it is male first, it is termed protandry or “first male.” Oysters, for example, are born male, grow, become female, and lay eggs; some oyster species change sex multiple times.
Chromosomal sex-determination systems
Mammalian sex is determined genetically by the presence of X and Y chromosomes. Individuals homozygous for X (XX) are female, while heterozygous individuals (XY) are male. The presence of a Y chromosome causes the development of male characteristics, while its absence results in the development of female characteristics. The XY system is also found in some insects and plants (Fig. 14.1).
Avian sex determination is dependent on the presence of Z and W chromosomes (Fig 14.1). Homozygous for Z (ZZ) results in a male, while heterozygous (ZW) results in a female. The W appears to be essential in determining the sex of the individual, similar to the Y chromosome in mammals. Some fish, crustaceans, insects (such as butterflies and moths), and reptiles use this system.

Sex determination was discovered in the mealworm by the American geneticist Nettie Stevens in 1903 (Fig 14.2).

X and Y sex chromosomes
The XX/XY sex-determination system is likely the most familiar, as it is found in humans. The XX/XY system is found in most other mammals, as well as some insects. In this system, most females have two of the same kind of sex chromosome (XX), while most males have two distinct sex chromosomes (XY). The X and Y sex chromosomes are different in shape and size from each other, unlike the rest of the chromosomes (autosomes), and are sometimes called allosomes. In some species, such as humans, organisms remain sex indifferent for a period of time after fertilization; in others, however, such as fruit flies, sexual differentiation occurs as soon as the egg is fertilized.
Some species, such as fruit flies, use the presence of two X chromosomes to determine femaleness, called the X-centered sex determination system. Species that use the number of Xs to determine sex are nonviable with an extra X chromosome.
In the XX/X0 sex chromosome system, a variant of the XY system, females have two copies of the sex chromosome (XX) but males have only one (X0). The 0 denotes the absence of a second sex chromosome. Generally in this method, the sex is determined by amount of genes expressed across the two chromosomes. This system is observed in a number of insects, including the grasshoppers and crickets of order Orthoptera and in cockroaches (order Blattodea). A small number of mammals also lack a Y chromosome. These include the Amami spiny rat (Tokudaia osimensis) and the Tokunoshima spiny rat (Tokudaia tokunoshimensis) and Sorex araneus, a shrew species. Transcaucasian mole voles (Ellobius lutescens) also have a form of XO determination, in which both sexes lack a second sex chromosome. This mechanism of sex determination is not currently well-understood.
ZW/ZZ sex chromosomes
The ZW sex-determination system is found in birds, some reptiles, and some insects and other organisms. The ZW sex-determination system is reversed compared to the XY system: females have two different kinds of chromosomes (ZW), and males have two of the same kind of chromosomes (ZZ). Like the X/Y system, there are variations on this system. For example, there are moths and butterflies that are ZW, but some have been found female with Z0, as well as female with ZZW. Also, while mammals deactivate one of their extra X chromosomes when female, it appears that in the case of Lepidoptera (butterflies and moths), the males produce double the normal amount of enzymes, due to having two Z chromosomes. Because the use of ZW sex determination is varied, it is still unknown how exactly most species determine their sex.
Despite the similarities between the ZW and XY systems, these sex chromosomes evolved separately. In the case of the chicken, their Z chromosome is more similar to humans’ autosome 9. The chicken’s Z chromosome also seems to be related to the X chromosome of the platypus. When a ZW species, such as the Komodo dragon, reproduces parthenogenetically (asexually), usually only males are produced. This is due to the fact that the haploid eggs double their chromosomes, resulting in ZZ or WW. The ZZ become males, but the WW are not viable and are not brought to term.
In both XY and ZW sex determination systems, the sex chromosome carrying the critical factors is often significantly smaller, carrying little more than the genes necessary for triggering the development of a given sex.
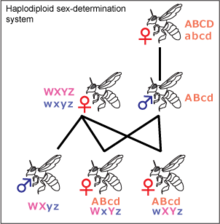
Haplodiploidy
Haplodiploidy is found in insects belonging to Hymenoptera, such as ants and bees. Sex determination is controlled by the zygosity of a complementary sex determiner (csd) locus. Unfertilized eggs develop into haploid individuals which have a single, hemizygous copy of the csd locus and are therefore males. Fertilized eggs develop into diploid individuals which, due to high variability in the csd locus, are generally heterozygous females. In rare instances diploid individuals may be homozygous, these develop into sterile males. The gene acting as a csd locus has been identified in the honeybee and several candidate genes have been proposed as a csd locus for other Hymenopterans. Most females in the Hymenoptera order can decide the sex of their offspring by holding received sperm in their spermatheca and either releasing it into their oviduct or not. This allows them to create more workers, depending on the status of the colony.
Environmental systems
The sex of some species is not determined by genetics, but by some aspect of the environment. Sex determination in some crocodiles and turtles, for example, is often dependent on the temperature during critical periods of egg development. This is referred to as environmental sex determination or, more specifically, as temperature-dependent sex determination. In many turtles, cooler temperatures during egg incubation produce males, while warm temperatures produce females. In some crocodiles, moderate temperatures produce males, while both warm and cool temperatures produce females.
Temperature-dependent sex determination
In some species of reptiles, including alligators, some turtles, and the tuatara, sex is determined by the temperature at which the egg is incubated during a temperature-sensitive period, called temperature-dependent sex determination (TSD). For some species with TSD, sex determination is achieved by exposure to hotter temperatures resulting in the offspring being one sex and cooler temperatures resulting in the other. This type of TSD is called Pattern I. For others species using TSD, it is exposure to temperatures on both extremes that results in offspring of one sex, and exposure to moderate temperatures that results in offspring of the opposite sex, called Pattern II TSD. The specific temperatures required to produce each sex are known as the female-promoting temperature and the male-promoting temperature. When the temperature stays near the threshold during the temperature sensitive period, the sex ratio is varied between the two sexes. Some species’ temperature standards are based on when a particular enzyme is created. Interestingly, the sex of some species, such as the Nile tilapia, Australian skink lizard, and Australian dragon lizard, is initially determined by chromosomes, but can later be changed by the temperature of incubation.
It is unknown how exactly temperature-dependent sex determination evolved. It could have evolved through certain sexes being more suited to certain areas that fit the temperature requirements. For example, a warmer area could be more suitable for nesting, so more females are produced to increase the amount that nest next season. Environmental sex determination preceded the genetically determined systems of birds and mammals; it is thought that a temperature-dependent amniote was the common ancestor of amniotes with sex chromosomes.
Other systems
There are other environmental sex determination systems including location-dependent determination systems as seen in the marine worm Bonellia viridis – larvae become males if they make physical contact with a female, and females if they end up on the bare sea floor. This is triggered by the presence of a chemical produced by the females, bonellin. Some species, such as some snails, practice sex change: adults start out male, then become female. In tropical clown fish, the dominant individual in a group becomes female while the other ones are male, and bluehead wrasses (Thalassoma bifasciatum) are the reverse. Some species, however, have no sex-determination system. Hermaphrodite species include the common earthworm and certain species of snails. A few species of fish, reptiles, and insects reproduce by parthenogenesis and are female altogether. There are some reptiles, such as the boa constrictor and Komodo dragon that can reproduce both sexually and asexually, depending on whether a mate is available.
Acknowledgements
Adapted from Boundless. (2023). General Biology. LibreTexts. Retrieved from https://bio.libretexts.org/Bookshelves/Introductory_and_General_Biology/Book%3A_General_Biology_(Boundless).
Adapted from https://en.wikipedia.org/wiki/Sex-determination_system
Ogilvie & Choquette (1981) Nettie Maria Stevens (1861-1912): Her Life and Contributions to Cytogenetics. Proceedings of the American Philosophical Society, 125(4), 292-311.

{kind=link}