27 Chapter 27: Animal Nutrition and Digestion
Mason Tedeschi and Joshua Reid
Learning Objectives
By the end of this section, students should be able to:
27.1 Be able to predict how different molecules (based on charge/polarity and size) would move across a membrane.
27.2 Be able to differentiate between intracellular and extracellular digestion and give examples of when you would see these.
27.3 Compare and contrast the processes of mechanical and chemical digestion.
27.4 Model the animal digestive, cardiovascular, or respiratory organ system, including the relationship and primary functions of its major organs.
27.5 Evaluate experimental data and results to determine whether claims about nutrition can be supported or not.
The plasma membrane, the cell membrane, has many functions, but the most basic one is to define the cell’s borders and keep the cell functional. The plasma membrane is selectively permeable. This means that the membrane allows some materials to freely enter or leave the cell, while other materials cannot move freely, but require a specialized structure, and occasionally, even energy investment for crossing.
Fluid Mosaic Model
Scientists identified the plasma membrane in the 1890s, and its chemical components in 1915. The principal components they identified were lipids and proteins. In 1935, Hugh Davson and James Danielli proposed the plasma membrane’s structure. This was the first model that others in the scientific community widely accepted. It was based on the plasma membrane’s “railroad track” appearance in early electron micrographs. Davson and Danielli theorized that the plasma membrane’s structure resembles a sandwich. They made the analogy of proteins to bread, and lipids to the filling. In the 1950s, advances in microscopy, notably transmission electron microscopy (TEM), allowed researchers to see that the plasma membrane’s core consisted of a double, rather than a single, layer. In 1972, S.J. Singer and Garth L. Nicolson proposed a new model that provides microscopic observations and better explains plasma membrane function.
The explanation, the fluid mosaic model, has evolved somewhat over time, but it still best accounts for plasma membrane structure and function as we now understand them. The fluid mosaic model describes the plasma membrane structure as a mosaic of components—including phospholipids, cholesterol, proteins, and carbohydrates—that gives the membrane a fluid character. Plasma membranes range from 5 to 10 nm in thickness. For comparison, human red blood cells, visible via light microscopy, are approximately 8 µm wide, or approximately 1,000 times wider than a plasma membrane. The membrane does look a bit like a sandwich (Figure 27.1).
A plasma membrane’s principal components are lipids (phospholipids and cholesterol), proteins, and carbohydrates attached to some of the lipids and proteins. A phospholipid is a molecule consisting of glycerol, two fatty acids, and a phosphate-linked head group. Cholesterol, another lipid comprised of four fused carbon rings, is situated alongside the phospholipids in the membrane’s core. The protein, lipid, and carbohydrate proportions in the plasma membrane vary with cell type, but for a typical human cell, protein accounts for about 50 percent of the composition by mass, lipids (of all types) account for about 40 percent, and carbohydrates comprise the remaining 10 percent. However, protein and lipid concentration varies with different cell membranes. For example, myelin, an outgrowth of specialized cells’ membrane that insulates the peripheral nerves’ axons, contains only 18 percent protein and 76 percent lipid. The mitochondrial inner membrane contains 76 percent protein and only 24 percent lipid. The plasma membrane of human red blood cells is 30 percent lipid. Carbohydrates are present only on the plasma membrane’s exterior surface and are attached to proteins, forming glycoproteins, or attached to lipids, forming glycolipids.
Phospholipids
The membrane’s main fabric comprises amphiphilic, phospholipid molecules. The hydrophilic or “water-loving” areas of these molecules (which look like a collection of balls in an artist’s rendition of the model) (Figure 27.1) are in contact with the aqueous fluid both inside and outside the cell. Hydrophobic, or water-hating molecules, tend to be non-polar. They interact with other non-polar molecules in chemical reactions, but generally do not interact with polar molecules. When placed in water, hydrophobic molecules tend to form a ball or cluster. The phospholipids’ hydrophilic regions form hydrogen bonds with water and other polar molecules on both the cell’s exterior and interior. Thus, the membrane surfaces that face the cell’s interior and exterior are hydrophilic. In contrast, the cell membrane’s interior is hydrophobic and will not interact with water. Therefore, phospholipids form an excellent two-layer cell membrane that separates fluid within the cell from the fluid outside the cell.
A phospholipid molecule (Figure 27.2) consists of a three-carbon glycerol backbone with two fatty acid molecules attached to carbons 1 and 2, and a phosphate-containing group attached to the third carbon. This arrangement gives the overall molecule a head area (the phosphate-containing group), which has a polar character or negative charge, and a tail area (the fatty acids), which has no charge. The head can form hydrogen bonds, but the tail cannot. Scientists call a molecule with a positively or negatively charged area and an uncharged, or non-polar, area amphiphilic or “dual-loving.”
This characteristic is vital to the plasma membrane’s structure because, in water, phospholipids arrange themselves with their hydrophobic tails facing each other and their hydrophilic heads facing out. In this way, they form a lipid bilayer—a double layered phospholipid barrier that separates the water and other materials on one side from the water and other materials on the other side. Phospholipids heated in an aqueous solution usually spontaneously form small spheres or droplets (micelles or liposomes), with their hydrophilic heads forming the exterior and their hydrophobic tails on the inside (Figure 27.3).
Proteins
Proteins comprise the plasma membranes’ second major component. Integral proteins, or integrins, as their name suggests, integrate completely into the membrane structure, and their hydrophobic membrane-spanning regions interact with the phospholipid bilayer’s hydrophobic region (Figure 27.1). Single-pass integral membrane proteins usually have a hydrophobic transmembrane segment that consists of 20–25 amino acids. Some span only part of the membrane—associating with a single layer—while others stretch from one side to the other, and are exposed on either side. Up to 12 single protein segments comprise some complex proteins, which are extensively folded and embedded in the membrane (Figure 27.4). This protein type has a hydrophilic region or regions, and one or several mildly hydrophobic regions. This arrangement of protein regions orients the protein alongside the phospholipids, with the protein’s hydrophobic region adjacent to the phospholipids’ tails and the protein’s hydrophilic region or regions protruding from the membrane and in contact with the cytosol or extracellular fluid.
Peripheral proteins are on the membranes’ exterior and interior surfaces, attached either to integral proteins or to phospholipids. Peripheral proteins, along with integral proteins, may serve as enzymes, as structural attachments for the cytoskeleton’s fibers, or as part of the cell’s recognition sites. Scientists sometimes refer to these as “cell-specific” proteins. The body recognizes its own proteins and attacks foreign proteins associated with invasive pathogens.
Carbohydrates
Carbohydrates are the third major plasma membrane component. They are always on the cells’ exterior surface and are bound either to proteins (forming glycoproteins) or to lipids (forming glycolipids) (Figure 27.1). These carbohydrate chains may consist of 2–60 monosaccharide units and can be either straight or branched. Along with peripheral proteins, carbohydrates form specialized sites on the cell surface that allow cells to recognize each other. These sites have unique patterns that allow for cell recognition, much the way that the facial features unique to each person allow individuals to recognize him or her. This recognition function is very important to cells, as it allows the immune system to differentiate between body cells (“self”) and foreign cells or tissues (“non-self”). Similar glycoprotein and glycolipid types are on the surfaces of viruses and may change frequently, preventing immune cells from recognizing and attacking them.
We collectively refer to these carbohydrates on the cell’s exterior surface—the carbohydrate components of both glycoproteins and glycolipids—as the glycocalyx (meaning “sugar coating”). The glycocalyx is highly hydrophilic and attracts large amounts of water to the cell’s surface. This aids in the cell’s interaction with its watery environment and in the cell’s ability to obtain substances dissolved in the water. As we discussed above, the glycocalyx is also important for cell identification, self/non-self determination, and embryonic development, and is used in cell to cell attachments to form tissues.
Plasma membranes must allow certain substances to enter and leave a cell, and prevent some harmful materials from entering and some essential materials from leaving. In other words, plasma membranes are selectively permeable (semipermeable)—they allow some substances to pass through, but not others. If they were to lose this selectivity, the cell would no longer be able to sustain itself, and it would be destroyed. Some cells require larger amounts of specific substances. They must have a way of obtaining these materials from extracellular fluids. This may happen passively, as certain materials move back and forth, or the cell may have special mechanisms that facilitate transport. Some materials are so important to a cell that it spends some of its energy, hydrolyzing adenosine triphosphate (ATP), to obtain these materials. Red blood cells use some of their energy doing just that. Most cells spend the majority of their energy to maintain an imbalance of sodium and potassium ions between the cell’s interior and exterior, as well as on protein synthesis.
The most direct forms of membrane transport are passive. Passive transport is a naturally occurring phenomenon and does not require the cell to exert any of its energy to accomplish the movement. In passive transport, substances move from an area of higher concentration to an area of lower concentration. A physical space in which there is a single substance concentration range has a concentration gradient.
Selective Permeability
Plasma membranes are asymmetric: the membrane’s interior is not identical to its exterior. There is a considerable difference between the array of phospholipids and proteins between the two leaflets that form a membrane. On the membrane’s interior, some proteins serve to anchor the membrane to cytoskeleton’s fibers. There are peripheral proteins on the membrane’s exterior that bind extracellular matrix elements. Carbohydrates, attached to lipids or proteins, are also on the plasma membrane’s exterior surface. These carbohydrate complexes help the cell bind required substances in the extracellular fluid. This adds considerably to plasma membrane’s selective nature (Figure 27.5).
Recall that plasma membranes are amphiphilic: They have hydrophilic and hydrophobic regions. This characteristic helps move some materials through the membrane and hinders the movement of others. Non-polar and lipid-soluble material with a low molecular weight can easily slip through the membrane’s hydrophobic lipid core. Substances such as the fat-soluble vitamins A, D, E, and K readily pass through the plasma membranes in the digestive tract and other tissues. Fat-soluble drugs and hormones also gain easy entry into cells and readily transport themselves into the body’s tissues and organs. Oxygen and carbon dioxide molecules have no charge and pass through membranes by simple diffusion.
Polar substances present problems for the membrane. While some polar molecules connect easily with the cell’s outside, they cannot readily pass through the plasma membrane’s lipid core. Additionally, while small ions could easily slip through the spaces in the membrane’s mosaic, their charge prevents them from doing so. Ions such as sodium, potassium, calcium, and chloride must have special means of penetrating plasma membranes. Simple sugars and amino acids also need the help of various transmembrane proteins (channels) to transport themselves across plasma membranes.
Diffusion
Diffusion is a passive process of transport. A single substance moves from a high concentration to a low concentration area until the concentration is equal across a space. You are familiar with diffusion of substances through the air. For example, think about someone opening a bottle of ammonia in a room filled with people. The ammonia gas is at its highest concentration in the bottle. Its lowest concentration is at the room’s edges. The ammonia vapor will diffuse, or spread away, from the bottle, and gradually, increasingly more people will smell the ammonia as it spreads. Materials move within the cell’s cytosol by diffusion, and certain materials move through the plasma membrane by diffusion (Figure 27.6). Diffusion expends no energy. On the contrary, concentration gradients are a form of potential energy, which dissipates as the gradient is eliminated.
Each separate substance in a medium, such as the extracellular fluid, has its own concentration gradient, independent of other materials’ concentration gradients. In addition, each substance will diffuse according to that gradient. Within a system, there will be different diffusion rates of various substances in the medium.
Facilitated transport
In facilitated transport, or facilitated diffusion, materials diffuse across the plasma membrane with the help of membrane proteins. A concentration gradient exists that would allow these materials to diffuse into the cell without expending cellular energy. However, these materials are polar molecule ions that the cell membrane’s hydrophobic parts repel. Facilitated transport proteins shield these materials from the membrane’s repulsive force, allowing them to diffuse into the cell.
The transported material first attaches to protein or glycoprotein receptors on the plasma membrane’s exterior surface. This allows removal of material from the extracellular fluid that the cell needs. The substances then pass to specific integral proteins that facilitate their passage. Some of these integral proteins are collections of beta-pleated sheets that form a pore or channel through the phospholipid bilayer. Others are carrier proteins which bind with the substance and aid its diffusion through the membrane.
Channels
The integral proteins involved in facilitated transport are transport proteins, and they function as either channels for the material or carriers. In both cases, they are transmembrane proteins. Channels are specific for the transported substance. Channel proteins have hydrophilic domains exposed to the intracellular and extracellular fluids. In addition, they have a hydrophilic channel through their core that provides a hydrated opening through the membrane layers (Figure 27.7). Passage through the channel allows polar compounds to avoid the plasma membrane’s nonpolar central layer that would otherwise slow or prevent their entry into the cell. Aquaporins are channel proteins that allow water to pass through the membrane at a very high rate.
Channel proteins are either open at all times or they are “gated,” which controls the channel’s opening. When a particular ion attaches to the channel protein it may control the opening, or other mechanisms or substances may be involved. In some tissues, sodium and chloride ions pass freely through open channels; whereas, in other tissues a gate must open to allow passage. An example of this occurs in the kidney, where there are both channel forms in different parts of the renal tubules. Cells involved in transmitting electrical impulses, such as nerve and muscle cells, have gated channels for sodium, potassium, and calcium in their membranes. Opening and closing these channels changes the relative concentrations on opposing sides of the membrane of these ions, resulting in facilitating electrical transmission along membranes (in the case of nerve cells) or in muscle contraction (in the case of muscle cells).
Carrier Proteins
Another type of protein embedded in the plasma membrane is a carrier protein. This aptly named protein binds a substance and, thus triggers a change of its own shape, moving the bound molecule from the cell’s outside to its interior (Figure 27.8); depending on the gradient, the material may move in the opposite direction. Carrier proteins are typically specific for a single substance. This selectivity adds to the plasma membrane’s overall selectivity. Scientists poorly understand the exact mechanism for the change of shape. Proteins can change shape when their hydrogen bonds are affected, but this may not fully explain this mechanism. Each carrier protein is specific to one substance, and there are a finite number of these proteins in any membrane. This can cause problems in transporting enough material for the cell to function properly. When all of the proteins are bound to their ligands, they are saturated and the rate of transport is at its maximum. Increasing the concentration gradient at this point will not result in an increased transport rate.
An example of this process occurs in the kidney. In one part, the kidney filters glucose, water, salts, ions, and amino acids that the body requires. This filtrate, which includes glucose, then reabsorbs in another part of the kidney. Because there are only a finite number of carrier proteins for glucose, if more glucose is present than the proteins can handle, the excess is not transported and the body excretes this through urine. In a diabetic individual, the term is “spilling glucose into the urine.” A different group of carrier proteins, glucose transport proteins, or GLUTs, are involved in transporting glucose and other hexose sugars through plasma membranes within the body.
Channel and carrier proteins transport material at different rates. Channel proteins transport much more quickly than carrier proteins. Channel proteins facilitate diffusion at a rate of tens of millions of molecules per second; whereas, carrier proteins work at a rate of a thousand to a million molecules per second.
Reading Question #1
Which process allows materials move down a concentration gradient and across the plasma membrane with the help of membrane proteins?
a. Simple diffusion
b. Facilitated diffusion
c. Active transport
d. Selective permeability
Invertebrate Digestive Systems
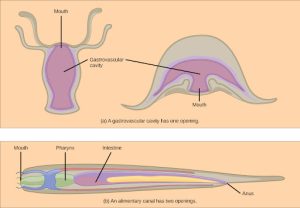
Animals have evolved different types of digestive systems to aid in the digestion of the different foods they consume. The simplest example is that of a gastrovascular cavity and is found in organisms with only one opening for digestion. Platyhelminthes (flatworms), Ctenophora (comb jellies), and Cnidaria (coral, jelly fish, and sea anemones) use this type of digestion. Gastrovascular cavities, as shown in Figure 27.9 a, are typically a blind tube or cavity with only one opening, the “mouth”, which also serves as an “anus”. Ingested material enters the mouth and passes through a hollow, tubular cavity. Cells within the cavity secrete digestive enzymes that breakdown the food. The food particles are engulfed by the cells lining the gastrovascular cavity.
The alimentary canal, shown in Figure 27.9b, is a more advanced system: it consists of one tube with a mouth at one end and an anus at the other. Earthworms are an example of an animal with an alimentary canal. Once the food is ingested through the mouth, it passes through the esophagus and is stored in an organ called the crop; then it passes into the gizzard where it is churned and digested. From the gizzard, the food passes through the intestine, the nutrients are absorbed, and the waste is eliminated as feces, called castings, through the anus.
Vertebrate Digestive Systems
Vertebrates have evolved more complex digestive systems to adapt to their dietary needs. Some animals have a single stomach, while others have multi-chambered stomachs. Birds have developed a digestive system adapted to eating unmasticated food.
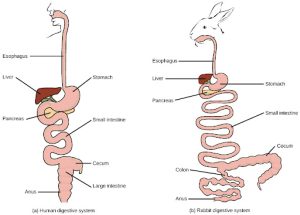
Monogastric: Single-chambered Stomach
As the word monogastric suggests, this type of digestive system consists of one (“mono”) stomach chamber (“gastric”). Humans and many animals have a monogastric digestive system as illustrated in Figure 27.10ab. The process of digestion begins with the mouth and the intake of food. The teeth play an important role in masticating (chewing) or physically breaking down food into smaller particles. The enzymes present in saliva also begin to chemically breakdown food. The esophagus is a long tube that connects the mouth to the stomach. Using peristalsis, or wave-like smooth muscle contractions, the muscles of the esophagus push the food towards the stomach. In order to speed up the actions of enzymes in the stomach, the stomach is an extremely acidic environment, with a pH between 1.5 and 2.5. The gastric juices, which include enzymes in the stomach, act on the food particles and continue the process of digestion. Further breakdown of food takes place in the small intestine where enzymes produced by the liver, the small intestine, and the pancreas continue the process of digestion. The nutrients are absorbed into the bloodstream across the epithelial cells lining the walls of the small intestines. The waste material travels on to the large intestine where water is absorbed and the drier waste material is compacted into feces; it is stored until it is excreted through the rectum.
Avian
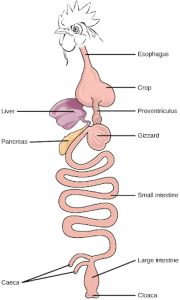
Birds face special challenges when it comes to obtaining nutrition from food. They do not have teeth and so their digestive system, shown in Figure 27.11 must be able to process un-masticated food. Birds have evolved a variety of beak types that reflect the vast variety in their diet, ranging from seeds and insects to fruits and nuts. Because most birds fly, their metabolic rates are high in order to efficiently process food and keep their body weight low. The stomach of birds has two chambers: the proventriculus, where gastric juices are produced to digest the food before it enters the stomach, and the gizzard, where the food is stored, soaked, and mechanically ground. The undigested material forms food pellets that are sometimes regurgitated. Most of the chemical digestion and absorption happens in the intestine and the waste is excreted through the cloaca.
Evolution Connection
Birds have a highly efficient, simplified digestive system. Recent fossil evidence has shown that the evolutionary divergence of birds from other land animals was characterized by streamlining and simplifying the digestive system. Unlike many other animals, birds do not have teeth to chew their food. In place of lips, they have sharp pointy beaks. The beak, lack of jaws, and the smaller tongue of the birds can be traced back to their dinosaur ancestors. The emergence of these changes seems to coincide with the inclusion of seeds in the bird diet. Seed-eating birds have beaks that are shaped for grabbing seeds and the two-compartment stomach allows for delegation of tasks. Since birds need to remain light in order to fly, their metabolic rates are very high, which means they digest their food very quickly and need to eat often. Contrast this with the ruminants, where the digestion of plant matter takes a very long time
Ruminants
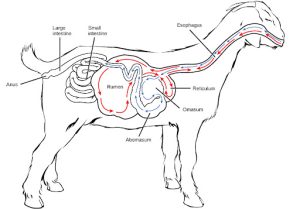
Ruminants are mainly herbivores like cows, sheep, and goats, whose entire diet consists of eating large amounts of roughage or fiber. They have evolved digestive systems that help them digest vast amounts of cellulose. An interesting feature of the ruminants’ mouth is that they do not have upper incisor teeth. They use their lower teeth, tongue and lips to tear and chew their food. From the mouth, the food travels to the esophagus and on to the stomach.
To help digest the large amount of plant material, the stomach of the ruminants is a multi-chambered organ, as illustrated in Figure 27.12. The four compartments of the stomach are called the rumen, reticulum, omasum, and abomasum. These chambers contain many microbes that breakdown cellulose and ferment ingested food. The abomasum is the “true” stomach and is the equivalent of the monogastric stomach chamber where gastric juices are secreted. The four-compartment gastric chamber provides larger space and the microbial support necessary to digest plant material in ruminants. The fermentation process produces large amounts of gas in the stomach chamber, which must be eliminated. As in other animals, the small intestine plays an important role in nutrient absorption, and the large intestine helps in the elimination of waste.
Pseudo-ruminants
Some animals, such as camels and alpacas, are pseudo-ruminants. They eat a lot of plant material and roughage. Digesting plant material is not easy because plant cell walls contain the polymeric sugar molecule cellulose. The digestive enzymes of these animals cannot breakdown cellulose, but microorganisms present in the digestive system can. Therefore, the digestive system must be able to handle large amounts of roughage and breakdown the cellulose. Pseudo-ruminants have a three-chamber stomach in the digestive system. However, their cecum—a pouched organ at the beginning of the large intestine containing many microorganisms that are necessary for the digestion of plant materials—is large and is the site where the roughage is fermented and digested. These animals do not have a rumen but have an omasum, abomasum, and reticulum.
Parts of the Digestive System
The vertebrate digestive system is designed to facilitate the transformation of food matter into the nutrient components that sustain organisms.
Oral Cavity
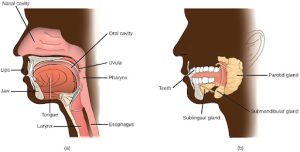
The oral cavity, or mouth, is the point of entry of food into the digestive system, illustrated in Figure 27.13. The food consumed is broken into smaller particles by mastication, the chewing action of the teeth. All mammals have teeth and can chew their food.
The extensive chemical process of digestion begins in the mouth. As food is being chewed, saliva, produced by the salivary glands, mixes with the food. Saliva is a watery substance produced in the mouths of many animals. There are three major glands that secrete saliva—the parotid, the submandibular, and the sublingual. Saliva contains mucus that moistens food and buffers the pH of the food. Saliva also contains immunoglobulins and lysozymes, which have antibacterial action to reduce tooth decay by inhibiting growth of some bacteria. Saliva also contains an enzyme called salivary amylase that begins the process of converting starches in the food into a disaccharide called maltose. Another enzyme called lipase is produced by the cells in the tongue. Lipases are a class of enzymes that can breakdown triglycerides. The lingual lipase begins the breakdown of fat components in the food. The chewing and wetting action provided by the teeth and saliva prepare the food into a mass called the bolus for swallowing. The tongue helps in swallowing—moving the bolus from the mouth into the pharynx. The pharynx opens to two passageways: the trachea, which leads to the lungs, and the esophagus, which leads to the stomach. The trachea has an opening called the glottis, which is covered by a cartilaginous flap called the epiglottis. When swallowing, the epiglottis closes the glottis and food passes into the esophagus and not the trachea. This arrangement allows food to be kept out of the trachea.
Reading Question #2
Chewing (masticating) is an important part of the digestion process, except in:
A. Rabbits
B. Goats
C. Birds
D. Humans
Esophagus
The esophagus is a tubular organ that connects the mouth to the stomach. The chewed and softened food passes through the esophagus after being swallowed. The smooth muscles of the esophagus undergo a series of wave like movements called peristalsis that push the food toward the stomach, as illustrated in Figure 27.14. The peristalsis wave is unidirectional—it moves food from the mouth to the stomach, and reverse movement is not possible. The peristaltic movement of the esophagus is an involuntary reflex; it takes place in response to the act of swallowing.
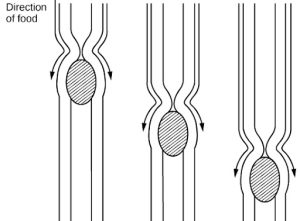
A ring-like muscle called a sphincter forms valves in the digestive system. The gastro-esophageal sphincter is located at the stomach end of the esophagus. In response to swallowing and the pressure exerted by the bolus of food, this sphincter opens, and the bolus enters the stomach. When there is no swallowing action, this sphincter is shut and prevents the contents of the stomach from traveling up the esophagus. Many animals have a true sphincter; however, in humans, there is no true sphincter, but the esophagus remains closed when there is no swallowing action. Acid reflux or “heartburn” occurs when the acidic digestive juices escape into the esophagus.
Stomach
A large part of digestion occurs in the stomach, shown in Figure 27.15. The stomach is a saclike organ that secretes gastric digestive juices. The pH in the stomach is between 1.5 and 2.5. This highly acidic environment is required for the chemical breakdown of food and the extraction of nutrients. When empty, the stomach is a rather small organ; however, it can expand to up to 20 times its resting size when filled with food. This characteristic is particularly useful for animals that need to eat when food is available.
The stomach is also the major site for protein digestion in animals other than ruminants. Protein digestion is mediated by an enzyme called pepsin in the stomach chamber. Pepsin is secreted by the chief cells in the stomach in an inactive form called pepsinogen. Pepsin breaks peptide bonds and cleaves proteins into smaller polypeptides; it also helps activate more pepsinogen, starting a positive feedback mechanism that generates more pepsin. Another cell type—parietal cells—secrete hydrogen and chloride ions, which combine in the lumen to form hydrochloric acid, the primary acidic component of the stomach juices. Hydrochloric acid helps to convert the inactive pepsinogen to pepsin. The highly acidic environment also kills many microorganisms in the food and, combined with the action of the enzyme pepsin, results in the hydrolysis of protein in the food. Chemical digestion is facilitated by the churning action of the stomach. Contraction and relaxation of smooth muscles mixes the stomach contents about every 20 minutes. The partially digested food and gastric juice mixture is called chyme. Chyme passes from the stomach to the small intestine. Further protein digestion takes place in the small intestine. Gastric emptying occurs within two to six hours after a meal. Only a small amount of chyme is released into the small intestine at a time. The movement of chyme from the stomach into the small intestine is regulated by the pyloric sphincter.
When digesting protein and some fats, the stomach lining must be protected from getting digested by pepsin. There are two points to consider when describing how the stomach lining is protected. First, as previously mentioned, the enzyme pepsin is synthesized in the inactive form. This protects the chief cells, because pepsinogen does not have the same enzyme functionality of pepsin. Second, the stomach has a thick mucus lining that protects the underlying tissue from the action of the digestive juices. When this mucus lining is ruptured, ulcers can form in the stomach. Ulcers are open wounds in or on an organ caused by bacteria (Helicobacter pylori) when the mucus lining is ruptured and fails to reform.
Small Intestine
Chyme moves from the stomach to the small intestine. The small intestine is the organ where the digestion of protein, fats, and carbohydrates is completed. The small intestine is a long tube-like organ with a highly folded surface containing finger-like projections called the villi. The apical surface of each villus has many microscopic projections called microvilli. These structures, illustrated in Figure 27.16, are lined with epithelial cells on the luminal side and allow for the nutrients to be absorbed from the digested food and absorbed into the bloodstream on the other side. The villi and microvilli, with their many folds, increase the surface area of the intestine and increase absorption efficiency of the nutrients. Absorbed nutrients in the blood are carried into the hepatic portal vein, which leads to the liver. There, the liver regulates the distribution of nutrients to the rest of the body and removes toxic substances, including drugs, alcohol, and some pathogens.
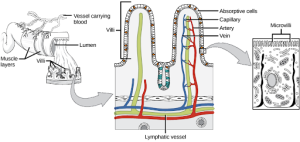
The human small intestine is over 6m long and is divided into three parts: the duodenum, the jejunum, and the ileum. The “C-shaped,” fixed part of the small intestine is called the duodenum and is shown in Figure 27.17. The duodenum is separated from the stomach by the pyloric sphincter which opens to allow chyme to move from the stomach to the duodenum. In the duodenum, chyme is mixed with pancreatic juices in an alkaline solution rich in bicarbonate that neutralizes the acidity of chyme and acts as a buffer. Pancreatic juices also contain several digestive enzymes. Digestive juices from the pancreas, liver, and gallbladder, as well as from gland cells of the intestinal wall itself, enter the duodenum. Bile is produced in the liver and stored and concentrated in the gallbladder. Bile contains bile salts which emulsify lipids while the pancreas produces enzymes that catabolize starches, disaccharides, proteins, and fats. These digestive juices breakdown the food particles in the chyme into glucose, triglycerides, and amino acids. The bulk of chemical digestion of food takes place in the duodenum. Absorption of fatty acids also takes place in the duodenum.
The second part of the small intestine is called the jejunum, shown in Figure 27.17. Here, hydrolysis of nutrients is continued while most of the carbohydrates and amino acids are absorbed through the intestinal lining. Some chemical digestion and the bulk of nutrient absorption occurs in the jejunum.
The ileum, also illustrated in Figure 27.17 is the last part of the small intestine and here the bile salts and vitamins are absorbed into the bloodstream. The undigested food is sent to the colon from the ileum via peristaltic movements of the muscle. The ileum ends and the large intestine begins at the ileocecal valve. The vermiform, “worm-like,” appendix is located at the ileocecal valve. The appendix of humans secretes no enzymes and has an insignificant role in immunity.
Large Intestine
The large intestine, illustrated in Figure 27.18, reabsorbs the water from the undigested food material and processes the waste material. The human large intestine is much smaller in length compared to the small intestine but larger in diameter. It has three parts: the cecum, the colon, and the rectum. The cecum joins the ileum to the colon and is the receiving pouch for the waste matter. The colon is home to many bacteria or “intestinal flora” that aid in the digestive processes. The colon can be divided into four regions, the ascending colon, the transverse colon, the descending colon, and the sigmoid colon. The main functions of the colon are to extract the water and mineral salts from undigested food, and to store waste material. Carnivorous mammals have a shorter large intestine compared to herbivorous mammals due to their diet.
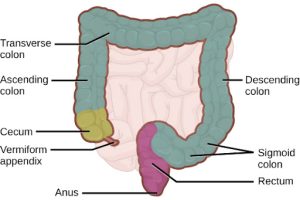
Rectum and Anus
The rectum is the terminal end of the large intestine. The primary role of the rectum is to store the feces until defecation. The feces are propelled using peristaltic movements during elimination. The anus is an opening at the far-end of the digestive tract and is the exit point for the waste material. Two sphincters between the rectum and anus control elimination: the inner sphincter is involuntary and the outer sphincter is voluntary.
Accessory Organs
The organs discussed above are the organs of the digestive tract through which food passes. Accessory organs are organs that add secretions (enzymes) that catabolize food into nutrients. Accessory organs include salivary glands, the liver, the pancreas, and the gallbladder. The liver, pancreas, and gallbladder are regulated by hormones in response to the food consumed.
The liver is the largest internal organ in humans and it plays a very important role in digestion of fats and detoxifying blood. The liver produces bile, a digestive juice that is required for the breakdown of fatty components of the food in the duodenum. The liver also processes the vitamins and fats and synthesizes many plasma proteins.
The pancreas is another important gland that secretes digestive juices. The chyme produced from the stomach is highly acidic in nature; the pancreatic juices contain high levels of bicarbonate, an alkali that neutralizes the acidic chyme. Additionally, the pancreatic juices contain a large variety of enzymes that are required for the digestion of protein and carbohydrates.
The gallbladder is a small organ that aids the liver by storing bile and concentrating bile salts. When chyme containing fatty acids enters the duodenum, the bile is secreted from the gallbladder into the duodenum.
Reading Question #3
Food gets to the stomach through the
A. Trachea
B. Esophagus
C. Gastrovascular cavity
D. Intestines
Ingestion
The large molecules found in intact food cannot pass through the cell membranes. Food needs to be broken into smaller particles so that animals can harness the nutrients and organic molecules. The first step in this process is ingestion. Ingestion is the process of taking in food through the mouth. In vertebrates, the teeth, saliva, and tongue play important roles in mastication (preparing the food into bolus). While the food is being mechanically broken down, the enzymes in saliva begin to chemically process the food as well. The combined action of these processes modifies the food from large particles to a soft mass that can be swallowed and can travel the length of the esophagus.
Digestion and Absorption
Digestion is the mechanical and chemical breakdown of food into small organic fragments. It is important to breakdown macromolecules into smaller fragments that are of suitable size for absorption across the digestive epithelium. Large, complex molecules of proteins, polysaccharides, and lipids must be reduced to simpler particles such as simple sugar before they can be absorbed by the digestive epithelial cells. Different organs play specific roles in the digestive process. The animal diet needs carbohydrates, protein, and fat, as well as vitamins and inorganic components for nutritional balance. How each of these components is digested is discussed in the following sections.
Carbohydrates
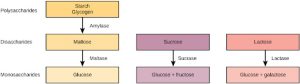
The digestion of carbohydrates begins in the mouth. The salivary enzyme amylase begins the breakdown of food starches into maltose, a disaccharide. As the bolus of food travels through the esophagus to the stomach, no significant digestion of carbohydrates takes place. The esophagus produces no digestive enzymes but does produce mucous for lubrication. The acidic environment in the stomach stops the action of the amylase enzyme.
The next step of carbohydrate digestion takes place in the duodenum. Recall that the chyme from the stomach enters the duodenum and mixes with the digestive secretion from the pancreas, liver, and gallbladder. Pancreatic juices also contain amylase, which continues the breakdown of starch and glycogen into maltose, a disaccharide. The disaccharides are broken down into monosaccharides by enzymes called maltases, sucrases, and lactases, which are also present in the brush border of the small intestinal wall. Maltase breaks down maltose into glucose. Other disaccharides, such as sucrose and lactose are broken down by sucrase and lactase, respectively. Sucrase breaks down sucrose (or “table sugar”) into glucose and fructose, and lactase breaks down lactose (or “milk sugar”) into glucose and galactose. The monosaccharides (glucose) thus produced are absorbed and then can be used in metabolic pathways to harness energy. The monosaccharides are transported across the intestinal epithelium into the bloodstream to be transported to the different cells in the body. The steps in carbohydrate digestion are summarized in Figure 27.19.
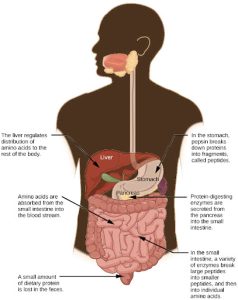
Protein
A large part of protein digestion takes place in the stomach. The enzyme pepsin plays an important role in the digestion of proteins by breaking down the intact protein to peptides, which are short chains of four to nine amino acids. In the duodenum, other enzymes—trypsin, elastase, and chymotrypsin—act on the peptides reducing them to smaller peptides. Trypsin elastase, carboxypeptidase, and chymotrypsin are produced by the pancreas and released into the duodenum where they act on the chyme. Further breakdown of peptides to single amino acids is aided by enzymes called peptidases (those that breakdown peptides). Specifically, carboxypeptidase, dipeptidase, and aminopeptidase play important roles in reducing the peptides to free amino acids. The amino acids are absorbed into the bloodstream through the small intestines. The steps in protein digestion are summarized in Figure 27.20.
Lipids
Lipid digestion begins in the stomach with the aid of lingual lipase and gastric lipase. However, the bulk of lipid digestion occurs in the small intestine due to pancreatic lipase. When chyme enters the duodenum, the hormonal responses trigger the release of bile, which is produced in the liver and stored in the gallbladder. Bile aids in the digestion of lipids, primarily triglycerides by emulsification. Emulsification is a process in which large lipid globules are broken down into several small lipid globules. These small globules are more widely distributed in the chyme rather than forming large aggregates. Lipids are hydrophobic substances: in the presence of water, they will aggregate to form globules to minimize exposure to water. Bile contains bile salts, which are amphipathic, meaning they contain hydrophobic and hydrophilic parts. Thus, the bile salts hydrophilic side can interface with water on one side and the hydrophobic side interfaces with lipids on the other. By doing so, bile salts emulsify large lipid globules into small lipid globules.
Why is emulsification important for digestion of lipids? Pancreatic juices contain enzymes called lipases (enzymes that breakdown lipids). If the lipid in the chyme aggregates into large globules, very little surface area of the lipids is available for the lipases to act on, leaving lipid digestion incomplete. By forming an emulsion, bile salts increase the available surface area of the lipids many fold. The pancreatic lipases can then act on the lipids more efficiently and digest them. Lipases breakdown the lipids into fatty acids and glycerides. These molecules can pass through the plasma membrane of the cell and enter the epithelial cells of the intestinal lining. The bile salts surround long-chain fatty acids and monoglycerides forming tiny spheres called micelles. The micelles move into the brush border of the small intestine absorptive cells where the long-chain fatty acids and monoglycerides diffuse out of the micelles into the absorptive cells leaving the micelles behind in the chyme. The long-chain fatty acids and monoglycerides recombine in the absorptive cells to form triglycerides, which aggregate into globules and become coated with proteins. These large spheres are called chylomicrons. Chylomicrons contain triglycerides, cholesterol, and other lipids and have proteins on their surface. The surface is also composed of the hydrophilic phosphate “heads” of phospholipids. Together, they enable the chylomicron to move in an aqueous environment without exposing the lipids to water. Chylomicrons leave the absorptive cells via exocytosis. Chylomicrons enter the lymphatic vessels, and then enter the blood in the subclavian vein.
Vitamins
Vitamins can be either water-soluble or lipid-soluble. Fat soluble vitamins are absorbed in the same manner as lipids. It is important to consume some amount of dietary lipid to aid the absorption of lipid-soluble vitamins. Water-soluble vitamins can be directly absorbed into the bloodstream from the intestine.
Reading Question #4
Emulsification is important for the digestion of:
A. Carbohydrates
B. Proteins
C. Lipids
D. All of the above
EliminationThe final step in digestion is the elimination of undigested food content and waste products. The undigested food material enters the colon, where most of the water is reabsorbed. Recall that the colon is also home to the microflora called “intestinal flora” that aid in the digestion process. The semi-solid waste is moved through the colon by peristaltic movements of the muscle and is stored in the rectum. As the rectum expands in response to storage of fecal matter, it triggers the neural signals required to set up the urge to eliminate. The solid waste is eliminated through the anus using peristaltic movements of the rectum.
Common Problems with Elimination
Diarrhea and constipation are some of the most common health concerns that affect digestion. Constipation is a condition where the feces are hardened because of excess water removal in the colon. In contrast, if enough water is not removed from the feces, it results in diarrhea. Many bacteria, including the ones that cause cholera, affect the proteins involved in water reabsorption in the colon and result in excessive diarrhea.
Emesis
Emesis, or vomiting, is elimination of food by forceful expulsion through the mouth. It is often in response to an irritant that affects the digestive tract, including but not limited to viruses, bacteria, emotions, sights, and food poisoning. This forceful expulsion of the food is due to the strong contractions produced by the stomach muscles. The process of emesis is regulated by the medulla.
Reading Question #5
In humans, mechanical digestion includes:
A. Mixing and propulsion
B. Breaking down carbohydrates and proteins
C. Bacteria aiding in the digestive process
D. Emesis
Adapted from Clark, M.A., Douglas, M., and Choi, J. (2018). Biology 2e. OpenStax. Retrieved from https://openstax.org/books/biology-2e/pages/1-introduction
